skip to main |
skip to sidebar
Davidson EA, Hart SC, Shanks CA, Firestone MK. 1991. Measuring gross nitrogen mineralization, immobilization, and nitrification by 15N isotopic pool dilution in intact soil cores. Journal of Soil Science 42: 335-349.These authors evaluated the use and limitations of the isotope-pool dilution technique when studying nitrogen dynamics in soil. Because addition of inorganic nitrogen compounds (NH4+, NO3-) can stimulate microbial activity in N-limited systems such as most soils, estimating the rate of these processes by tracking 15N through a system will almost certainly overestimate these rates. The isotope-pool dilution method, on the other hand, measures the dilution of enrichment in the nitrogen pool at the end of a particular process, relying on the assumption that additional product of metabolism will have negligible effects on the magnitude of that metabolism. In this study, immobilization of nitrogen was the main focus of investigation, comparing 15N isotope dilution in pools of either 15NH4+ or 15NO3-.There are three key assumptions for the isotope-pool dilution method in this context. 1. Microorganisms do not discriminate between 15N and 14N; 2. rates of processes measured remain constant over the incubation period; 3. 15N assimilated during the incubation period is not remineralized. Previously, these assumptions had been evaluated for well-mixed soils, but not for unmixed field-collected soil samples. While fractionation by biological processes certainly does result in discrimination between isotopes of nitrogen, it is of negligible importance when injected solutions are very highly enriched and incubation periods are relatively short; in this case, injections were more than 90% 15N and incubations ran for 24 hours. Rates of measured processes will change if the population and / or activity of microorganisms changes, but again, over a 24-hour incubation period under controlled conditions this is unlikely. Highly enriched injections allow the use of small injection volumes, limiting the impact of nutrient enrichment. These authors were able to measure the remineralization of immobilized 15N, and estimated that between 1.0 and 1.6% of injected 15NH4+ appeared in the 15NO3- pool after 24 hours; they consider this an insignificant amount, but caution that longer incubations would almost certainly result in much more problematic amounts of remineralization.This paper is clearly a major part of the basis of the project I am currently engaged in with Katherine from our 2009 field season at Alexandra Fjord. I probably should have read this paper long ago. The three main conclusions stated by these authors at the end of their paper I think can be quoted verbatim as justification for both why I (should have earlier) read this paper, and as a reminder to myself to include this paper in the methods & materials section of the eventual manuscript."Three points should be considered when applying the isotope dilution method.
1. Accurate estimation of both 14N and 15N initial pool sizes is important. Abiotic consumption of label, such as by clay fixation, can cause significant errors. A subset of intact cores may need to be destructively sampled directly after adding 15N for estimation of initial pool sizes.
2. Homogeneity of 15N enrichment throughout a soil sample is not possible, and perfectly uniform distribution of added label is not necessary. However, significant errors can arise from a bias in 15N distribution that is concurrent with a non-random
distribution of microbial processes. Distribution of label should, therefore, be as uniform as possible.
3. In situ gross immobilization rates may be overestimated by isotope dilution methods and underestimated by chloroform fumigation methods, depending on which (if any) kN factor is applied to the latter. Gross mineralization and gross nitrification estimates from isotope dilution are more reliable because these rates should not be affected by addition of 15N label in the form of the process products."
Miller MN, Zebarth BJ, Dandie CE, Burton DL, Goyer C, Trevors JT. 2008. Crop residue influence on denitrification, N2O emissions and denitrifier community abundance in soil. Soil Biology & Biochemistry 40: 2553-2562.These authors conducted a factorial experiment using packed soil cores to examine the influence of varying levels of available carbon and nitrogen on the process of denitrification. The treatments consisted of addition of glucose at three levels and KNO3 at four levels in experiment 1, and additions of either red clover or barley straw crop residues with or without additional KNO3. They measured soil chemistry, including extractable organic carbon and NO3- concentration based on K2SO4 extractions, as well as N2O production, the molar ratio of N2O (N2O : (N2O + N2)), and a handful of bacterial genes by qPCR.The experimental setup was very similar to what we used in SLSC 802 (Special Topics) in the fall of 2010; cylindrical soil cores with gas-exchange holes in the sides were filled with soil at 1 g cm-3 bulk density and a water content of 70% and placed in 1 L canning jars with lids fitted with a perforable septum for gas sampling. One important difference between this experiment and what we have done is that in this experiment, gas measurements were of total cumulative gas production, whereas we flushed each jar with ambient air after each sampling event. Presumably this difference will have important effects on the formation of anaerobic conditions and microbial consumption of N2O previously produced under less anoxic conditions.Not surprisingly, minimal denitrification activity was found in treatments without added NO3-. Starting NO3- concentrations were 3 mg NO3--N kg-1 soil, and fell in all treatments without added NO3- to less than 1 mg NO3--N kg-1. Once this supply of readily available nitrate was used, it appears the bacteria ceased denitrification activity, or at least it was reduced.The red clover had a much lower C:N ratio than the barley straw, 13:1 and 45:1, respectively, and more labile carbon. This difference appears to have driven the observed difference in denitrification activity, in a manner that reflects the results of the simple-C-source experiment 1. In general, more labile C and more available N leads to stronger denitrification activity and greater production of N2O; in sealed jars such as these, strong respiration under these conditions leads to anaerobic conditions and a fall in the molar ratio of N2O as nosZ-equipped microbes consume N2O as a terminal electron acceptor.Extractable organic carbon (EOC) was a relatively poor predictor of denitrification, compared to respiration as measured by CO2 production. EOC is a measure of the instantaneous size of the pool of labile C, while respiration represents carbon that has already passed through a microbe’s metabolism. The distinction here may be between two different pools of carbon, as well as between an instantaneous snapshot measure and a series of measurements readily convertible to an estimate of the rate of a process.In conclusion, these authors reiterate their finding that available C and available N (especially as NO3-) are strong predictors of denitrifying activity, across a range of C and N sources. I read this paper for the class SLSC 802 in the fall of 2010, but the portions describing denitrification physiology and especially the qPCR information will be generally useful to my other projects.
Delgado JA, Del Grosso SJ, Ogle SM. 2010. 15N isotopic crop residue cycling studies and modeling suggest that IPCC methodologies to assess residue contributions to N2O-N emissions should be reevaluated. Nutrient Cycling in Agroecosystems 86: 383-390.These authors reanalyzed two recent reviews of measuring nitrous oxide emissions from agricultural systems and used a model to simulate N2O emissions and NO3 leaching associated with cropping practices in Colorado and Iowa. In general, use of crop residue instead of or in addition to synthetic fertilizers significantly altered patterns of N loss, whereas IPCC recommendations assume no difference between these N sources in regards to N2O emissions. Microbial immobilization of nitrogen, particularly associated with residues with high C/N ratios, is a major factor in these differences, and these authors provide supporting arguments for their suggestion of revisions to IPCC recommendations and modeling.
Huang Y, Zou J, Zheng X, Wang Y, Xu X. 2004. Nitrous oxide emissions as influenced by amendment of plant residues with different C:N ratios. Soil Biology & Biochemistry 36: 973-981.These authors examined the role of residue quality, in the form of C:N ratio and a range of crop residues, on N2O emissions from soils. They also measured CO2 emissions, and found strong correlations between organic-matter decomposition and respiration, and nitrogen cycling.Gas fluxes of CO2 and N2O were highly correlated across all incubations. To ensure only respiration-derived CO2 was measured, the CO2 released by urea breakdown in urea-treated treatments was calculated and subtracted; respiration in the urea-only treatment was similar to that in the untreated controls. Both gas emissions were negatively correlated with residue C:N ratios. Finally, residue C:N ratios were negatively correlated with dissolved organic carbon concentrations. Overall, higher C:N ratios in residues seem to result in slow decomposition of mainly recalcitrant organic matter, and low CO2 and N2O emissions. Addition of urea in conjunction with crop residues produces a range of N2O emissions depending on the C:N ratio of the residues.This short paper may serve as a model for the work I will be doing in the special topics class in soil science, fall 2010.
Trinsoutrot I, Recous S, Mary B, Nicolardot B. 2000. C and N fluxes of decomposing 13C and 15N Brassica napus L.: effects of residue composition and N content. Soil Biology and Biochemistry 32: 1717-1730.These authors studied the decomposition process by soil microorganisms when isotope-labelled crop residues were added to soil. The crop used, oilseed rape Brassica napus (also known as canola) varies its nitrogen content of tissues, and the C:N ratio, depending on levels of N inputs by fertilization. This allows variation in input organic matter quality by manipulation of growing conditions; in this experiment, both carbon and nitrogen inputs to the plant included stable-isotope labels, in the form of 13C-CO2 and 15N-KNO3. Plant residues were added to soils and incubated for 168 days.Initial C:N ratio and especially the labile-C fraction of organic-matter inputs are major controls of both the rate of decomposition and fate of matter through the system. Additionally, temperature, particle size of residues, and water content in the soil also strongly influence decomposition processes. Here, N mineralization (the formation of NO3- and NH4+ pools in the soil from organic-N precursors) occurred in two phases. In the early phase, up to about 3 weeks, the N cycle resulted in net mineralization. Later, mineral N pools were depleted and N was immobilized, that is, incorporated into the tissues of microbial cells.Carbon dioxide release during the experiment occurred through two pathways. The more direct route was rapid mineralization of organic matter, which I interpret as non-incorporation of organic matter by microbes, consuming such material but metabolizing it rapidly through respiration. The second, presumably slower route was through metabolization of material after incorporation into cells through respiration. Either way, the ultimate fate of much of the organic-C in the residues was release as CO2.Differences in the N-content of residues affected decomposition rates early in the experiment, but by about 4 months the differences between high-N and low-N residues had evened out. Only a small fraction of labelled N from residues ended up in soil mineral-N pools; the majority was either immobilized into microbial cells or remained in recalcitrant organic matter fractions. Immobilization of unlabeled, SOM-derived N was enhanced by the addition of C through a substitution effect.These authors conclude that 15N labelling was fraught with difficulties, and both under- and overestimated some pools and processes. However, the use of their model, named NCSOIL, improved their ability to trace the fate of added material through the system. This paper represents a study similar in some ways to our planned course activity in the special topics in soil science course, fall 2010.
Smith KA, Ball T, Conen F, Dobbie KE, Massheder J, Rey A. 2003. Exchange of greenhouse gases between soil and atmosphere: interactions of soil physical factors and biological processes. European Journal of Soil Science 54:779-791.These authors present a broad review paper of the role of soil physical factors, mainly temperature and water-filled-pore-space, in controlling soil emissions of the greenhouse gases CO2, CH4, and N2O. The paper’s goal is stated to be to expose a variety of researchers to the links between soil physics and soil biology, as well as the importance of these fields to current research in many disciplines on global warming.All three gases are produced and consumed in soil primarily by microorganisms, which respond to variation in soil physical parameters in different ways. In general, both temperature and WFPS impact GHG production. Higher temperatures almost always result in increased production of gases, though the Q10 values (measuring the magnitude of response to a change of 10º of temperature) vary widely in the literature for all three gases. The effect of WFPS is different, involving upper and lower bounds, though in the middle range increasing WFPS generally promotes increased gas production. Microbes are limited in their tolerance of dry conditions, such that gas production falls rapidly below some critical WFPS value; for CO2 this threshold is near 20%. WFPS is also indirectly important, through its effects on soil diffusivity. Higher WFPS as well as higher bulk density are associated with lessened CH4 oxidation, due to reduced diffusivity of both CH4 and atmospheric O2. Very high WFPS values are associated with reduction of N2O to N2, partly by limiting O2 supplies and creating larger anaerobic microsites, and partly by preventing the escape of N2O gas into rapid-diffusion pathways; it is trapped in the vicinity of microbes capable of using it as an electron acceptor.There are other factors controlling net GHG emissions, such as the relationship between plant productivity and water table position, which will change the relationship between rates of soil organic matter oxidation to CO2 and the removal of CO2 from the atmosphere by plants; trees in particular can lower local water tables, increasing SOM oxidation while simultaneously consuming more CO2 than the previous wetland vegetation community. I read this paper on the suggestion of my coworkers in the special topics class of fall 2010, but it applies well to the general area of my research. The reference list includes multiple interesting papers addressing particular specialties within this large topic.
Pennock DJ, Zebarth BJ, De Jong E. 1987. Landform classification and soil distribution in hummocky terrain, Saskatchewan, Canada. Geoderma 40: 297-315.These authors present a method of analysing irregular terrain for the purposes of examining important soil parameters such as soil depth and soil hydrology. Three variables (profile curvature, plan curvature, and gradient) can be calculated from a matrix of elevation data; taken together for a given point or area, these variables can be used to classify an area into one of seven landforms. These landforms are level (for summits or bottom lands), shoulders, backslopes, and footslopes; the three non-level forms come in “divergent” (convex plan) and “convergent” (concave plan) varieties. Shoulders have convex profiles, backslopes have flat profiles (i.e. constant gradient when looking up or down the slope), and footslopes are concave. Divergent landforms shed water laterally; convergent landforms tend to collect water. Where water collects and is moving slowly, rates of infiltration will be highest, and erosion will tend to deposit, rather than remove, material at these places.The variables required to calculate profile and plan curvature and gradient are relatively easy to calculate from a matrix of elevation data, using an interpolating topographical software package and some differential calculus. The seven categories of slope elements can be estimated in the field by eye, making for a useful method for field studies.This paper was on the recommended reading list for SLSC 834; in addition, the course instructor is Dr. Pennock, lead author of this study. The study site, near Hafford, Saskatchewan, is perhaps 1 to 1.5 hours drive away from Saskatoon, suggesting this area may be the destination of one of the day trips scheduled for the week of August 30, 2010.
Hayashi M, van der Kamp G, Rudolph DL. 1998. Water and solute transfer between a prairie wetland and adjacent uplands, 1. Water balance. Journal of Hydrology 207: 42-55.These authors report water balance data and analysis in the first of a pair of papers about the hydrology of wetlands in the prairie pothole region of central Saskatchewan. The second paper concerns dissolved material movements in the same example study system.Through a series of measurements of water and water-related properties of soil as well as precipitation, total water balance and total water movements within the wetland were estimated. Direct precipitation and evaporation accounts for a small fraction of the total volume of water entering and leaving the pond in the middle of the hectare-scale wetland and surrounding catchment of uplands. Most of the water entering the pond is in the form of snowmelt during spring, and most of the water leaving the pond is in the form of horizontal groundwater flow, relatively shallowly (i.e. above a redox discontinuity at about 5-6m depth), driven strongly by evapotranspiration through the “willow ring” surrounding the pond and through crops (principally wheat during this experiment) on the uplands. Only a small amount of water is exported down to the aquifer; of a total input of 780mm into the pond, only about 3mm goes to recharging the underlying aquifer. Figure 3 provides definitions of the terminology, and Figure 12 the estimates of water balance.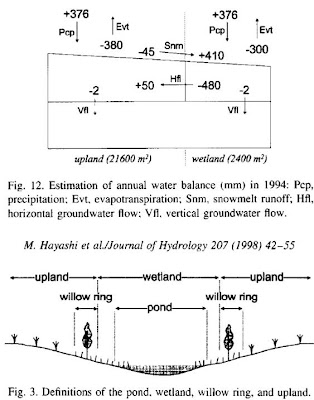 This paper was on the suggested reading list for SLSC 834 in August 2010.
This paper was on the suggested reading list for SLSC 834 in August 2010.
Christiansen EA. 1979. The Wisconsinan deglaciation of southern Saskatchewan and adjacent areas. Canadian Journal of Earth Science 16: 913-938. This author describes in considerable detail the process of deglaciation that occurred at the end of the last glacial maximum from about 17 000 years ago, as it occurs to Saskatchewan. Major geological features and patterns of melt-water drainage led to the inclusion of nearby parts of Alberta, Manitoba, Montana, and North Dakota in the analysis. Patterns during the deglaciation were identified by glacial landforms, many of which are apparent only from aerial photographs, with sample analysis in the lab including radio-carbon dating.The period from 17 000 years to 10 000 years ago is divided into 9 phases corresponding to periods of rapid glacial retreat or temporary stasis or glacial advance. By 10 000 years ago the ice sheet that had covered nearly the entire province had retreated towards Hudson Bay and covered only a small part of the north of Saskatchewan. Glacial lakes formed from meltwater and from water flowing from the west (presumably sourced from glaciers in the Rocky mountains), sometimes reaching enormous sizes; these lakes were bordered by the glacier’s edge, and connected to each other via spillways that often carved large channels from the plains; the east-west valleys of southern Saskatchewan such as the Qu’Appelle valley are the remains of such spillways. Modern river systems such as the Saskatchewan River and the Churchill River formed during the glacial retreat, occupying low areas and spillway remnants.The rate of deglaciation varied considerably over the studied 7 000 years, generally accelerating from about 150 m / yr to around 275 m / yr, though with frequent pauses, occasional re-advances, and variation across the glacial edge. The evidence from glacial lake-edge movements and depth patterns suggests the ice sheet melted fastest initially, but with the slowest retreat at that time indicating the sheet first thinned, and then retreated, especially around newly-uncovered Nunataks where the underlying land formed highlands.This paper was on the suggested reading list for the course SLSC 834 in August 2010, but it is also personally interesting in describing Pleistocene events in areas I visit during Sunday drives across the center of the province.
Wilkinson MT, Richards PJ, Humphreys GS. 2009. Breaking ground: Pedological, geological, and ecological implications of soil bioturbation. Earth-Science Reviews 97: 257-272.These authors describe the process of bioturbation in soils, and the roles of various major groups of organisms, especially animals, responsible for bioturbation. Through mixing and surface-mounding, bioturbation sorts, buries, and transports soil and soil components such as organic matter. Bioturbation also interacts with abiotic processes, particularly those involving surface water inputs and movements, to create net effects on soil movement and sorting.There is a great deal of detail in the description of various sub-processes and rates, as well as some urging for more detailed studies of these phenomena. This paper is required reading for the course SLSC 834, in August 2010.
Johnson DL, Domier JEJ, Johnson DN. 2005. Reflections on the nature of soil and its biomantle. Annals of the Association of American Geographers 95: 11-31.These authors advocate a new paradigm to underlie studies of soil science and related fields, based on increased recognition of processes occurring in and responsible for the production of the “biomantle”, the upper layer of soil composed of and formed by the actions of organisms. Many of these processes are based on movements of soil and soil components, and so are dominated by animals, especially active burrowers that are responsible for large vertical movements of soil in some environments. The biomantle is defined as being composed of “biofabric”, or materials that owe their existence to the actions of organisms, from the bodies of these organisms themselves, to the materials released by the organisms, to the minerals created by biological processes, to the voids created by their movements and the gases filling those voids released by their metabolisms.These authors trace their ideas from the writings of Darwin, particularly his final work involving the activity of worms in “vegetable mould”, a late-18th century term for what we now call soil. A biomantle layer, residing chiefly in the A horizon (or topsoil) can be more easily recognized in some landscapes than others. Humid tropical soils especially may show very thick and distinct biomantles, in two layers. The upper, thicker layer is composed of relatively fine materials, resting on a basal layer of coarser material; this basal layer is referred to here as the stonelayer. Below the stonelayer is non-biomantle, typically a B horizon (or subsoil). The hypothesized process creating this two-layer biomantle is the action of “conveyor belt” animals, especially termites that carry small particles upwards but are unable to move larger stones, thus eventually sorting the soil mineral material. In other soils, such as loess-derived sandy soils without a large component of gravel and larger stones, such a two-layer biomantle may not form, or may be very weakly developed and difficult to identify as such. Nonetheless, bioturbation activity by burrowing animals is usually apparent, for example in the form of “krotovina”, in-filled animal burrows.Besides advocating for a view of soils and their processes with an animal-based, biomantle point of view, these authors spend some time dismissing subaqueous soils (e.g. marine sediments) as simplistic, uncomplicated places lacking many of the key (and very complex) processes that occur in subaerial soils. Their list of such processes near the end of the paper, taken as a kind of justification for their uncited and unsupported dismissal of subaqueous soils, is composed entirely of those processes relating to changing water amounts in terrestrial soils, such as groundwater flow and wetting and drying events. I found their argument unconvincing, as they do not describe any aqueous-only processes such as changes in dissolved-O2 concentrations or the sorting action of water currents, and their blithe disregard for marine biodiversity in statements about how much more diverse the life in terrestrial soils must be, is the proverbial icing on the insult cake. Johnson et al.: please cite some evidence to support such sweeping generalizations.
Stark JM, Hart SC. 1996. Diffusion technique for preparing salt solutions, Kjeldahl digests, and persulfate digests for nitrogen-15 analysis. Soil Science Society of America Journal 60: 1846-1855.These authors evaluated the recently-developed Teflon-encased-acid-trap technique for collecting nitrogen from water samples for analysis, especially stable isotope (15N) analysis. Earlier acid trap techniques, which rely on the chemistry of ammonium/ammonia in solutions of varying pH, were based on suspended small vials containing acid solutions above a larger volume of sample solution at high pH. Ammonium present in the sample is converted to ammonia by high pH, and escapes solution into the gas phase. It is captured in acid solution; in a closed container with a separate acidic region, ammonium will migrate from the alkaline sample to the acid trap. Suspended trap designs are vulnerable to a number of problems, including vulnerability of the acid trap to contamination by alkaline solution and subsequent loss of captured ammonium. Teflon will pass gases but not liquids, and therefore provides an ideal barrier between the sample and the acid trap that will permit ammonia to enter the trap.These authors conducted a set of six experiments to evaluate the utility of Teflon acid traps. First, blanks were evaluated, to determine the sensitivity of the technique to nitrogen derived from contamination in the various materials of the experiment (filter paper, Teflon, plastic bottles, etc.). There was some nitrogen detectable from these non-sample sources, but a correction factor could be constructed based on including a few blanks in sample sets. The second experiment examined the recovery of ammonium from 2M KCl solutions. These solutions are very similar to the solutions extracted from soils of Alexandra Fjord in 2009. As might be expected, incubation time correlated with recovery of ammonium; in addition, Teflon traps performed better over shorter incubations (up to about 6 days) than did suspended traps, though the difference disappeared at the longest (8 days) incubations. The third experiment was of greatest interest to me. They examined the open-bottle time necessary to eliminate residual ammonium from samples used in experiment 2, and the incubation conditions to collect NO3- -derived nitrogen using Devarda’s alloy to reduce NO3- to NH4+. Residual ammonium left in solution at the end of the first reaction would contaminate NO3- examination and lead to overestimation of NO3- contents; this is especially true when the two forms of nitrogen have been previously enriched with 15N to different degrees. There is a trade-off between open bottle time (ranging in these experiments from one to five days) and recovery of NO3- from samples; long open-bottle times appear to lead to an unknown chemical change in samples, which these authors speculate, may involve sorption of atmospheric CO2. Long ammonium-collection times (i.e. experiment 2) do tend to collect the great majority of ammonium present, recovering on average more than 97% of ammonium from known-concentration samples. This suggests to me a long and potentially nitrate-losing open-bottle incubation is not necessary if ammonium collection has proceeded for at least six days. Agitation of samples, in these experiments often once per day, resulted in a strong improvement in nitrogen capture over at least the shorter (24-96 hour) incubations; our use of continuous agitation for seven days therefore seems likely to capture nearly all ammonium present in samples.Experiments 4 through 6 were of little interest to me, as I am unfamiliar with Kjeldahl digests and persulfate digests and do not plan to use these techniques in my studies. From what I could glean based on my limited knowledge, the Teflon-acid-trap technique appears to work well with Kjeldahl digests and persulfate digests.Overall, the Teflon-acid-trap technique performed very well, and had significant ease-of-use and contamination-avoiding advantages over earlier methods. One caution put forth by these authors is regarding the H2 gas evolved during Devarda’s alloy incubations; strong build up of gas can cause leaks, potentially allowing NH3 to escape and leading to underestimates of sample nitrate contents. They suggest storing bottles upside-down during such incubations, as leaks will then include sample solution, and leaking bottles can be easily identified. Complete recovery of nitrogen was not possible on a routine basis using this technique, but 15N isotope ratios do not rely on 100% recovery if ammonium and / or nitrate concentrations have been measured by an independent method, such as the widely-used colorimetric techniques. The two isotopes of nitrogen do not migrate differently between alkaline sample and acid trap, thus even at relatively inefficient recovery rates, isotopic ratios are maintained. Blanks are important, but in general this technique is robust, reliable, and easy to conduct.
Palmer K, Drake HL, Horn MA. 2010. Association of novel and highly diverse acid-tolerant denitrifiers with N2O fluxes of an acidic fen. Applied and Enironmental Microbiology 76: 1125-1134.These authors examined soils from an acidic fen in southern Germany, and discovered novel denitrifiers that are apparently adapted to local conditions and contribute to the cycling of nitrogen within the fen. Methods employed included measurement of soil parameters, microcosms to examine denitrification rates (both total denitrification and net production / consumption of N2O), cell counts of cultured organisms, and phylogenetic analysis using narG and nosZ sequences and RFLP.
Palmer K, Drake HL, Horn MA. 2009. Genome-derived criteria for assigning environmental narG and nosZ sequences to operational taxonomic units of nitrate reducers. Applied and Environmental Microbiology 75: 5170-5174.These authors compared the sequences of narG and nosZ genes to corresponding sequences of 16s rRNA genes, using in-silico analysis of sequences downloaded from GenBank. While similarities above 97% are commonly used for species- or genus-level taxonomic delineation for 16s sequences, this analysis found much lower threshold similarities for such delineation using the structural genes. This paper is confusing to me. One part of the text appears to contradict itself, when the authors state that the Nar operon in Pseudomonas stutzeri A1501 is putatively alien in origin (i.e. recent horizontal transfer), then go on to state in the same paragraph that it appears unlikely that the Nar operon was horizontally transferred in any species. I may just be misunderstanding the meaning of the term “putatively alien” in regards to a bacterial gene sequence.A greater puzzle is presented by the list of nosZ sequences. These authors downloaded 85 such sequences, where my own attempts to extract data from GenBank resulted in only 42 unique nosZ sequences. The list in a supplementary table includes several cases of multiple accessions of the same species but of different PD. The paper these clusters of PD-sequences are derived from is Dandie et al. (2007); a quick scan of this paper did not reveal what the distinction “PD” indicates.
Tuomivirta TT, Yrjälä K, Frize H. 2009. Quantitative PCR of pmoA using a novel reverse primer correlates with potential methane oxidation in Finnish Fen. Research in Microbiology 160: 751-756.These authors developed novel primers for studying the methanotrophs of Finnish peatland fens. The existing primer pairs widely used to study methanotrophs, based on the gene pmoA, did not consistently amplify useful products in PCR using template DNA from these fens.The novel primer pair was tested on 114 samples from two Finnish fens, representing the northern and southern peatlands of Finland. While A189f/A682r failed to provide useful products, the new primer A6821r in conjunction with A189f did produce strong bands. In qPCR, these primers produced results correlated with measured methane oxidation potential, further supporting their utility in these systems.
Schmidt SK, Reed SC, Nemergut DR, Grandy AS, Cleveland CC, Weintraub MN, Hill AW, Costellow EK, Meyer AF, Neff JC, Martin AM. 2008. The earliest stages of ecosystem succession in high-elevation (5000 metres above sea level), recently deglaciated soils. Proceedings of the Royal Society of London, Series B 275: 2793-2802.These authors describe the microbial community and soil parameters of a chronosequence at the foreground of a receding glacier high in the Peruvian Andes. From a combination of aerial photography and previous work at this site, a series of sites of soils of increasing ages from zero to 79 years old was established. No surface plants, even lichens, are present on any of these new soils, and soil nutrient levels (carbon, nitrogen) are very low; the only organisms present are microorganisms.Two previous hypotheses had been proposed to explain the dynamics of very early primary succession on new soil. Organic matter has been observed to accumulate slowly in new soils; the source of this material is either aeolian deposits (i.e. wind-borne plant detritus and pollen) or in-situ fixation of CO2 and N2. These are not mutually exclusive hypotheses, but the relative contributions of each are explored in this study.The methods used here cover an extensive list of soil parameters. Three sets of soil samples were collected: for microbiological analysis, N-fixation measurement, and all other chemical analyses. The other chemical analyses include photosynthetic pigment extraction, soil total and mineral nitrogen, pyrolysis for identifying sources of carbon compounds (i.e. microbial-autotroph, microbial-heterotroph, plant), enzyme assays for common and informative microbial enzymes, and soil stability analysis of the resistance of these new soils to erosion forces such as water runoff.These authors focused on the cyanobacterial fraction of the microbial community in this study; some details of other components of the biota are described in an earlier paper, Nemergut et al. (2007). Cyanobacteria are autotrophs also capable of fixing atmospheric nitrogen, thus they are ideal primary colonizers of new soil as they require little more than a source of moisture and air. Analysis of the community included the use of the P-test (Martin 2002); note that as in Nemergut et al. (2007), he is one of the authors of this study. The analytical approach is very similar to that employed in the earlier study, with a comparison of discovered sequences to published sequences from around the world. In this study, cyanobacterial sequences from zero and 4-year-old soils were similar to sequences from an extremely broad sample of habitats, including Antarctic lake ice, marine subseafloor sediments, urban aerosols, forest soils, and oil-polluted soils. The soil chronosequence showed a clear pattern of stages of primary succession at every level of analysis. The soil microbial community became both more abundant and more diverse through time, soil nutrients increased, the chemical environment included increasing amounts and diversity of complex organic molecules, key enzyme pathways became established, and soil stability increased as soils aged. N-fixation showed a peak, with increasing N-fixation activity from the zero to 4-year-old soils (by two orders of magnitude), then declining by about half in the 79-year-old soils. This mirrors and precedes a widely-observed pattern in plant primary succession, in which nitrogen-fixing plants are among the first colonizers, but decline in abundance at later stages of succession. Enzyme and organic molecule patterns were consistent with a total absence of heterotrophs in the extremely young soils, increasing occurrence of organisms capable of decomposing plant matter in the 4-year-old soils, and a molecular ecology qualitatively similar to a mature plant-associated soil in the 79-year-old soil. The list of procedures and level of detail of analysis in this paper is impressive. Many, though certainly not all, of these techniques will be models for my own work, especially in the summer of 2010. The molecular-diversity techniques pioneered by Martin (Martin 2002, Nemergut et al. 2007, this paper) as well as the techniques of analyzing soil pigments and soil nutrients are all very interesting.
Sørensen LI, Holmstrup M, Maraldo K, Christensen S, Christensen B. 2006. Soil fauna communities and microbial respiration in high Arctic tundra soils at Zackenberg, Northeast Greenland. Polar Biology 29: 189-195.These authors sampled soil animals from three sites at Zackenberg station, Greenland, over three days in mid-summer. Two of the sites were considered mesic heath, with a mix of Cassiope tetragona and other High Arctic species of plants, while the third site was dominated by Dryas spp. and was considered dry heath; snow melts from the dry heath up to 20 days earlier than from the mesic heaths. Soil samples ranging down to about 6cm depth were collected, stored at 5ºC for up to two weeks, and analyzed by a range of methods in the laboratories in Europe.Different groups of soil animals were extracted by varying methods. Soil microarthropods, a diverse group dominated by Collebola and Acari, were extracted by modified MacFadyen funnels into Benzoic acid. Enchytraeids and dipteran larvae were extracted in Baermann wet funnels with heating of the samples, into tap water. Protozoa were washed from soils in water and grown on media plates in the dark at 10ºC. Nematodes were collected by the Blender-Cotton wool method of Schouten and Arp (1991). Soil microbial respiration was measured in serum bottles, with the CO2 concentration in the headspace measured at zero, 5 and 25 hours, with a fully factorial design of nutrient amendments of C, N, and P. Soil pH and soil organic matter content, but not moisture content or other nutrient concentrations were determined using methods not clearly described, though presumably these procedures were similar to standard methods.Once abundance and biomass data was collected, comparisons between plots were made using multivariate analysis and a software package named PRIMER 5.0. My understanding is the species counts were (log+1) transformed to reduce the influence of very abundant species, then analyzed using an approach similar to Principal Components Analysis. The result of this analysis was a clear difference between the dry heath and the two mesic heaths, while the two mesic heaths were not different from each other in parameter-space. A Bray-Curtis similarity matrix was also involved, though I’m not certain I understand how.Different taxonomic groups were identified to different taxonomic levels; 19 species of Collembola and 7 species of Enchytraeids were found, for example, but Acari were identified to suborder (Cryptostigmata (oribatids), Prostigmata, Mesostigmata) and nematodes and protozoans were counted at those high taxonomic levels. While the two mesic heath sites were only marginally significantly different from each other, there was a clear increase in abundances in the dry heath site. For collembola at least, the dry heath site was also dominated by two highly abundant species, which differed from the majority of species in the mesic sites by being unpigmented and associated with sub-surface, rather than soil-surface, regions in the soil. The higher abundance of probably bacteria-eating nematodes at the dry heath strongly suggests higher turnover of microorganisms as well as generally higher biological activity from the higher populations of most soil animals.These authors suggest higher organic matter decomposition rates at the dry heath, which seems reasonable given the higher animal populations there. However, their attribution of higher soil pH there to higher respiration levels seems like more of a stretch, absent supporting mineralogical and soil-nutrient data.This paper provides an excellent example of the data that can be collected and analyzed from a brief but intensive study of soil invertebrates at a High Arctic site. In addition, meaningful information about differences in biodiversity between locations can be derived from studies of organisms not identified to fine taxonomic levels.
O’Neill KP, Godwin HW, Jiménez-Esquilín AE, Battigelli JP. 2010. Reducing the dimensionality of soil microinvertebrate community datasets using Indicator Species Analysis: Implications for ecosystem monitoring and soil management. Soil Biology & Biochemistry 42: 145-154.These authors used a dataset of soil microarthropods to evaluate a method for identifying indicator species for ecosystem monitoring. The method centres on the Indicator Value (IV) of a species, a number that integrates the degree of uniqueness to a place of a species and the abundance of that species within a given habitat. A high IV value indicates both high information content and a high probability of being sampled. The IV is apparently robust to differences in site number and species absolute abundances, and provides a single value for evaluating observed or expected changes in an ecosystem. Indicator species, furthermore, integrate habitat conditions over their lifespans, in contrast to measures of chemical and physical parameters that capture a snapshot of an ecosystem.The basic evaluation approach here was to identify indicator species along a clear environmental gradient from meadow to forest in West Virginia. The habitat was divided into three zones, with an edge patch between the open meadows and closed-canopy forest. Near-surface soil cores were collected from each zone every month from April 2004 to April 2005 (n = 180), using the top of the mineral soil as the reference depth. Microarthropods were extracted in a modified Macfadyen funnel with a strong and increasing temperature gradient, into 70% ethanol.Diversity measures, including Simpson’s and Shannon indices, were based on counts of individuals identified to family level (suborder for Acari). Differences between sites were analyzed by 2-way repeated measures ANOVA and Principle Components Analysis, after rare taxa (those that occurred in less than 10% of samples) were removed; rare taxa are extremely unlikely to be identified as indicator species.Calculating IV for each taxon, regardless of the taxonomic resolution, provides large advantages in labour time and taxonomic expertise, as many microfauna are very difficult to identify to genus or species. These authors state that enumeration of a single sample required more than 1 hour of a trained taxonomist’s time. In studies such as this one, there are further advantages of IV associated with its robustness in the face of many zero measurements (i.e. taxa absent from samples) and the general messiness of these kinds of datasets. However, the ISA approach is intended for 2-stage studies, where an intensive initial survey identifies indicator species (taxa), and later long-term monitoring ignores other species. For studies specificially designed to address biodiversity, such as my own, excluding rare taxa would not be beneficial, and there may be no easy escape from time-consuming morphotaxa sorting.I have spoken with Dr. Battigelli, the trained taxonomist in this study. He has indicated that while this IV-based approach may not be appropriate for my own work, it nonetheless demonstrates the types of analyses that can be conducted with soil invertebrates identified to middle taxonomic levels. He has assured me I could probably be trained to identify Collembola to Family and Acari to Suborder in a matter of a few days, and he would be interested in futher studies of collected soil invertebrates based on interesting patterns that emerge at these taxonomic levels.
Harding RJ, Gryning S-E, Halldin S, Lloyd CR. 2001. Progress in understanding of land surface/atmosphere exchanges at high latitudes. Theoretical and Applied Climatology 70: 5-18.These authors review and discuss the implications of studies based in two international projects in northern Europe. WINTEX was a large study examining the effects of snow cover and long nights in winter on high-latitude ground-atmosphere exchange processes, while LAPP was an independent but complementary study examining most of the same processes in a range of high latitude sites during spring and summer.Snow cover plays a major role in Arctic exchange processes. The high albedo of snow reflects much of the incident solar radiation, and insulates the frozen ground below, prolonging the period of snow cover to upwards of 9 months in the year in many places. Where vegetation is tall, such as in the boreal forest, the low solar angle reduces the effective net albedo of the landscape, allowing sunlight to warm the dark trees and speed springtime melting. This study mentions the importance of snow-surface aerodynamics, though it appears there is little solid information on this complex topic.Snow melt is the major hydrological event of the year in much of the Arctic. The combination of frozen soils, very low evaporation rates, and often flat terrain means much of the Arctic is very wet or saturated while annual precipitation rates are consistent with arid or semi-arid conditions. These areas are the classic tundra systems, with abundant shallow lakes and ponds and very wet high-organic soils.Differences in snow-surface dynamics and the timing of snowmelt create an extremely heterogeneous landscape, particularly in the vicinity of the northern treeline. There are often very large temperature and air-flow differences between patches of trees and adjacent lakes or clearings, which greatly complicate attempts to model the carbon dioxide emissions (for example) of such areas. Much of this paper is a series of evaluations of some of the models that have been applied to this region. In general, more sophisticated models that can take some of the extreme variability into account perform better than models that cannot account for differences in snow depth or insulating properties. However, this paper makes it clear that current modelling efforts still leave much to be desired in terms of predicting Arctic heat budgets and biological responses.Water storage is also very difficult to model, and has large and variable impacts on other parts of the system. There appears to be large and unpredictable year-to-year variation in water storage and transport at the scale of catchments and basins, and the importance of soil water in controlling biological processes such as the decomposition of organic matter is large. Runoff matters, even on very gentle slopes.This paper provides a useful overview of large-scale processes and attempts to understand these processes in the Arctic.
Martin KJ, Rygiewicz PT. 2005. Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiology 5:28.These authors designed new primers for PCR and related molecular biology investigations of soil fungi, especially mycorrhizae. These are very diverse organisms, and, because of the commonalities between fungal and plant DNA, studies of fungal samples closely entwined with plant tissue as in root-associated mycorrhizae can be very complex. The new primers were designed to produce a range of PCR products suitable for techniques such as qPCR, Length-Heterogeneity PCR (LH-PCR) and T-RFLP analyses. The primers as a suite were designed around a nested approach, with new outer primers amplifying a long DNA sequence of approximately 1000bp, and later primer pairs amplifying regions within that long sequence. Most inner primer pairs generate products of approximately 500bp length. These authors also used a different DNA extraction method, based on xanthogenate and Tween (X/T) that involves little to no tissue grinding, compared to the standard method based on CTAB. The X/T method preferentially extracts fungal DNA from cells on the outside of particles, for example fungal cells not penetrating plant roots. This reduces the amount of plant DNA and associated plant-derived compounds in resulting extracts. Combining the two techniques may allow for some interesting studies of fungal micro-ecology.This paper’s novel primers should be more specific and more useful to my own qPCR studies of Arctic soil microbes, including soil fungi. The methods section detailing some of the decisions made and considerations involved in primer design will also be useful.
Philippot L, Piutti S, Martin-Laurent F, Hallet S, Germon JC. 2002. Molecular analysis of the nitrate-reducing community from unplanted and maize-planted soils. Applied and Environmental Microbiology 68: 6121-6128.These authors applied molecular techniques including PCR, RFLP, and sequencing to the study of soil bacteria relevant to crops. Dissimilatory nitrate reduction, the process that converts NO3- to NO2-, is widespread in prokaryotes, with the activity described in alpha, beta, and gamma Proteobacteria, gram-positive bacteria, and some archaea. There are two described enzymes that catalyze the reaction and provide energy to the organism; these authors focused on the membrane-bound protein, specifically one subunit that includes a distinctive set of components. Their approach was to design primers for a well-conserved region of the gene narG that amplify a 650bp region, and then subject the PCR product to cloning, RFLP analysis, and sequencing. Community structure and diversity was compared between pots planted with maize versus unplanted controls. Maize (Zea mays) is a plant that facilitates gas diffusion in its roots under oxygen-stress soil conditions; this creates an aerobic region in the rhizosphere distinct from anaerobic conditions further from roots. While diversity, as measured by standard indices, did not differ between planted and unplanted soils, the structure of the communities did change, with numerous RFLP phylotypes found in only one or the other treatment. This suggests a role of rhizosphere conditions, likely involving both oxygen and root exudates, in selecting for particular groups of microorganisms.Nitrate reduction occurs primarily or possibly only under aerobic conditions. The microbial cell gains energy from dissimilatory reduction of nitrate, and if it occurs in the rhizosphere, the plant may gain a readily-accessible form of nitrogen in the form of nitrite. Denitrification, the process that shuttles nitrogen atoms from nitrite to gaseous forms such as N2O or N2, can occur under a range of oxygen conditions, including aerobic, thus denitrifiers in the rhizosphere may compete with plant roots for nitrite. The fate of nitrite produced by dissimilatory nitrate reduction can also be to ammonium, though this appears to be rare in soil and more common in vertebrate guts and digested sludge, two environments typically lacking in oxygen.This paper provides some molecular tools for my own studies of nitrogen dynamics in soils, especially the sequences of the degenerate primers. In addition, it provides some clarification of parts of the remarkably complex soil-nitrogen cycle.
Siciliano SD, Ma W, Powell S. 2007. Evaluation of quantitative polymerase chain reaction to assess nosZ gene prevalence in mixed microbial communities. Canadian Journal of Microbiology 53: 636-642.These authors examined the usefulness of qPCR in studying populations of soil bacteria, especially denitrifiers using the gene nosZ that codes for nitrous oxide reductase. This enzyme catalyzes the final reaction in the process of denitrification, converting N2O to N2. Normally, it is expressed only in severely anaerobic conditions, as it allows the use of N2O as the terminal electron acceptor during metabolism.There are a number of factors that control the efficiency of PCR in quantitative PCR applications. The efficiency is a major component of the calculations that allow qPCR to estimate gene copy numbers in samples and thus to be used to examine population dynamics of non-culturable microorganisms from environmental samples. Of particular importance is consistency of efficiency between the amplification of the standard DNA template and the amplification of all templates in the unknown samples. Variation between the standard and the unknowns can lead to severe under- or over-estimation of target populations, while variation in efficiency between different templates within the unknown samples can lead to misestimations of relative proportions of organisms. These authors evaluated the efficiency of qPCR in a range of experimental templates, and in a range of combinations simulating mixed populations. Little variance in efficiency was found, and this variance was not associated with genetic distance from a reference organism. The experimental design did not allow a direct examination of the influence of the geographical differences in the sources of the test sequences (Arctic, temperate-grassland, Antarctic), but this lack of association with the reference organism does indicate low or no variation among PCR efficiencies associated with some other variable.The influence of varying PCR efficiencies among templates within a sample becomes less severe as the number of different templates rises. In a typical soil sample with perhaps 1000 different templates, no one template can utterly dominate amplification by outcompeting for primers, thus the resulting mix of amplicons at the end of 40 rounds of PCR will most likely be representative of the population mixture in the environment.This paper is of obvious high utility to my own work, not least because the individual machine used to perform qPCR is the same individual machine that I will be using. For this and other reasons, this paper was suggested to me, repeatedly. Future reference to this paper, when I am developing my methods and when I am writing up the next paper or two, seems likely.
Dandie CE, Miller MN, Burton DL, Zebarth BJ, Trevors JT, Goyer C. 2007. Nitric oxide reductase-targeted real-time PCR quantification of denitrifier populations in soil. Applied and Environmental Microbiology 73: 4250-4258.These authors examined the responses of two major components of the denitrifying bacteria fraction of soil bacteria to the addition of labile carbon (glucose) under denitrifying conditions. Denitrification is presented as a four-step process, with enzymes responsible for shuttling nitrate to N2 via nitrite, nitric oxide, and nitrous oxide. In this study, one of the enzymes responsible for the reduction of NO to N2O, cNOR, was examined using primers optimized for two different groups of denitrifying bacteria. This gene is found only in denitrifiers, unlike another enzyme, qNOR, found in many microorganisms and associated with detoxification, rather than utilization, of dangerous nitric oxide.Primers for qPCR are presented in a table. Specific primers for the two variants of cNOR were developed in this study for use with SYBR green-based qPCR. 16s rRNA sequences were also studied, to examine the total population of soil bacteria; for these qPCR reactions, the TaqMan primers-plus-probe system was used, based on oligonucleotides published by Suzuki et al. 2000. Two experiments were carried out. In the first, a preliminary experiment to establish the utility of qPCR in this area was based on inoculating soils with cultures of bacteria of known cell density, followed by qPCR evaluation of those soils. Under most conditions qPCR performed well, though at low cell densities of some genera of bacteria the signal was not distinguishable from the background noise also associated with sterilized soil. The second experiment forms the main body of work of this paper, and is an examination of the population dynamics of soil bacteria, divided into the hierarchical categories “denitrifiers” and “all bacteria”, under denitrifying conditions and with varying levels of added labile carbon in the form of glucose solutions in distilled water. In the second experiment, soil nitrate was maintained at a high level, to ensure sufficient raw material for detectable denitrification activity. As N2O accumulation was one of the measures of activity, nitrous oxide reductase activity that would reduce N2O to N2 was inhibited by maintaining an atmosphere of 10% acetylene in culture jars. Soils were maintained at 70% WFPS to encourage denitrification.Total microbial biomass was also measured, using the CHCl3 fumigation-extraction technique. While cNOR sequences are almost certainly restricted to one copy per genome, 16s rRNA sequences may range in copy number up to 15 per genome, thus estimates of bacterial populations by qPCR of 16s may have a large error associated with it. Fumigation-extraction captures all carbon associated with cells, thus contributions by archaea and fungi will not be found by molecular methods such as 16s qPCR that are specific to bacteria. However, in this study, estimates of total bacterial population by the two methods were well correlated, with r2 = 0.69.Denitrification occurred in this study. Soils treated with additional glucose showed greater depletion of nitrate, as expected when denitrifiers increase their activity in response to a food supply and conditions already favour denitrification. These authors provide two possible mechanisms, non-mutually-exclusive, that could lead to increased denitrification activity under added glucose. First, the population of denitrifiers could expand, through both additional cell replication and activation of dormant cells. This would increase the proportion of the bacterial population composed of denitrifiers. Second, the total population of soil organisms could increase, leading to increased respiration, a decrease in oxygenation, and establishment of anaerobic conditions more favourable for denitrification. This would not necessarily change the proportion of the population composed of denitrifiers. In this study, denitrifiers increased their proportion of the population as measured by comparative qPCR from less than 1% to about 2.4% of cell numbers.This change in population components is central to the approach using qPCR advocated in this paper. As these authors state:“Although absolute numbers may not be achievable, gross differences and changes in population size are still detectable. The differences observed between the two denitrifier populations studied are then real differences in the responses of these populations to the conditions tested.”
This general approach of examining relative changes in populations is applicable to a very wide array of studies of environmental microbiology, including my own planned studies in which the environmental factor under examination is biogeographical (i.e. latitude) and the functional diversity response is in terms of greenhouse gas cycling."This paper is of great value to my studies. The qPCR methods are directly applicable, for example the primers presented here will be useful if I decide to examine multiple components of the denitrification pathway. The approach, as described above, is also useful. And the reference list is composed almost entirely of papers I am surprised I have not yet found in my literature searches.
Liptzin D. 2006. A banded vegetation pattern in a High Arctic community on Axel Heiberg Island, Nunavut, Canada. Arctic, Antarctic, and Alpine Research 38: 216-223.This author attempted to explain the observation of banded vegetation on a slope that lacked the usual factors that generate such patterns. In temperate and tropical locations, banded vegetation, also known as “tiger stripes”, forms on shallow slopes in dry areas with a consistent direction of water flow. Plants at a position on the slope increase water retention and facilitate further colonization by plants. Similarly, some locations experience consistent wind direction carrying sea spray that kills trees at some positions. In cold environments, patterned ground from cryoturbation on shallow slopes can also lead to banded vegetation. However, the study site in this paper lacks all of these features, including cryoturbation despite the presence of permafrost within 50cm at most locations.Some aspect of soil properties is the obvious explanatory hypothesis, which this author explores after describing the transects measuring plant diversity and the soil pits used to examine soil properties. In general, features that would normally be expected to influence plant diversity and abundance such as soil moisture or exchangeable cation levels, had no significant impact in the various statistical tests employed in this study. However, soil type did have some effect, as a few species of plants were found only on sandy soil, and nitrogen levels were negatively correlated with species richness.The discussion section of this paper is an excellent example of a chain of logical reasoning working through a series of potential explanations. While this paper is interesting, it’s only relevant to my own studies in a narrow area around potential starting points in looking for explanations for whatever patterns I may find in my biogeography studies in 2010. However, this paper seems remarkably suitable as an introduction to the basics of modern soil science research, and may be relevant to my not-quite-mothballed interest in an undergraduate course about the current state of the scientific literature.
Klotz MG, Stein LY. 2008. Nitrifier genomics and evolution of the nitrogen cycle. FEMS Microbiology Letters 278: 146-156.These authors review the role of nitrifying microorganisms in the current nitrogen cycle, and their evolution and the emergence of biological nitrogen cycles in early Earth history. The current global nitrogen cycle has changed considerably in the past decades, due to the large increase in nitrogen in the cycle due to human activities. The early-Earth nitrogen cycle was probably mostly driven by abiotic processes. After the development of an oxygen-rich atmosphere, nitrogen cycling was almost entirely biotic, with most key processes driven largely or entirely by bacteria and archaea. In the last few decades, the anthropogenic abiotic processes of fertilizer production and fossil-fuel combustion combined with increased cultivation of N2-fixing crops, has transformed the global nitrogen cycle.As presented in this paper, there are two lobes to the global nitrogen cycle. N2 gas in the atmosphere is fixed to NH3, by nitrogenase in bacteria and archaea, by the Haber-Bosch industrial process, and (in small quantities) by hydrothermal vents. The process of nitrification converts this ammonia to nitrite/nitrate. Nitrite/nitrate are returned to the atmosphere as N2 through denitrification, with production of N2O under weakly anaerobic conditions. The other lobe of the cycle is a “short circuit” that avoids the atmospheric N2 pool and cycles nitrite/nitrate back to ammonia through the processes of ammonification and through production and decomposition of organic matter containing nitrogen. There is another, minor short circuit, as “anammox”, anaerobic ammonia oxidation, returns ammonia to N2 directly.This “mini review” focuses on the nitrification portion of the cycle. The first step, oxidation of ammonia to hydroxylamine (NH2OH) is carried out by ammonia oxidizing bacteria, abbreviated AOB. There are many acronyms in this paper, reflecting the many acronyms in the existing literature regarding global biogeochemical cycles. NOB are nitrite oxidizing bacteria, and they take the oxidized products of AOB, especially nitrite through to nitrate. Anammox bacteria, on the other hand, may run the same net process of NH3 to NO3- directly, without collaboration with other cells.The discussion of the plausible evolutionary scenarios in this paper is interesting but not particularly relevant to my current research. This discussion focuses on the relative timing of major events, such as the emergence of nitrification, complete and incomplete denitrification, an oxygenated atmosphere, and nitrogen fixation. These factors interact with each other, creating conditions favourable or not to the evolution of each other and of possible detail shifts within.The description of the role of hydroxylamine produced by early nitrifiers in stimulating evolution of metabolic pathways responsible for its detoxification initially reads as speculation, but a long and detailed description of the ways in which the various components of those metabolic detox enzymes and pathways function provides plenty of support for the arguments. One aspect of this discussion is that some enzymes are currently misclassified, and that very similar enzymes in different organisms have different names reflecting different ultimate functions rather than the usual (and preferred) enzyme naming scheme that reflects proximate function.In the discussion concerned with anthropogenic climate change and nitrogen dynamics, especially in soils and ocean waters, interactions with methane are briefly considered. This is based on the observation that methanotrophs are often also ammonia-oxidizers, operating under a budget of consumption of both molecules that shifts as ammonia from fertilizer is added to the system. Of greatest relevance to my current work is the section describing the gene ncyA. This encodes an enzyme (nitrosocyanin) involved in the pathway from ammonia to nitrite, and has only been found in AOB to date, as opposed to NOB, anammox, or heterotrophs; it seems to be involved in the chemistry of obligate chemolithotrophy as expressed by AOB. It seems likely the enzyme binds and reduces NO, a highly toxic intermediate in ammonia oxidation. The regulatory region adjacent to the gene is also suggestive of roles in this metabolic pathway, and regulation is linked to concentration of various nitrogen-with-oxygen compounds.This review is very useful to my current research. This paper and the major references in it will be key to constructing a diagram of the complex nitrogen transformations occurring in soils, which will allow targeted hypothesis generation and testing regarding the communities and processes in the soils I am studying.
Li X-R, Du B, Fu H-X, Wang R-F, Shi J-H, Wang Y, Jetten MSM, Quan ZX. 2009. The bacterial diversity in an anaerobic ammonium-oxidizing (anammox) reactor community. Systematic and Applied Microbiology 32: 278-289.These authors studied the bacterial community that developed inside a bioreactor running on sewage sludge under anaerobic conditions. Like Lim et al. (2008), the main focus of this study was in the applications of ammonia-oxidizing bacteria (AOB) to water treatment facilities. The expected chemistry of anaerobic ammonia oxidation catalyzed by microorganisms (“anammox”) includes the use of nitrite as the electron acceptor in a near-one-to-one ratio with the consumption of ammonia or ammonium. The energy derived from this process is used by the cell to fix CO2, thus making these organisms autotrophs. This alters the underlying stoichiometry slightly, as some nitrite is diverted to CO2 fixation rather than ammonia oxidation.The study of anaerobic AOB is still quite new, with the five described genera of such organisms all named with “Candidatus” prefixes, indicating recent species descriptions. All are in one group (taxonomic level unknown), the Brocadiales, within the phylum Planctomycetes. Aerobic AOB are in other groups, and include some species within the genus Nitrosomonas in the Beta-Proteobacteria that are capable of limited ammonia-oxidizing activity under anaerobic conditions, and can apparently survive long periods without oxygen.These authors did not develop novel primers for PCR or qPCR in this study. Instead, they used published primer sets; I gather they did not use the TaqMan double-dye system for qPCR, as no mention of probes is made. The target genomic sequences were portions of the 16s rRNA gene, using E. coli as a standard. Oddly, the overall procedure included normal PCR, followed by cloning and insertion into plasmids, followed by qPCR of plasmid DNA containing the 16s sequences. It is unclear to me exactly why this was done, though later in the paper there are a few sequence-based phylogenetic trees that might have been based on sequences derived from this cloning procedure. In any case, the qPCR did provide informative results regarding the composition of bacterial groups within the reactor.Of the sequences identified, the great majority were unlike cultivated organisms, highlighting the utility of these techniques in studying environmental samples. AnAOB produced approximately 16% of sequences, with aerobic AOB less than 1%. Non-AOB in three phyla constituted the majority of sequences, including 38% Chlorobi, 21% Chloroflexi, and 7% Bacteriodetes. These are filamentous heterotrophic bacteria, and appear to be closely associated with the granules that formed in the reactor solution after a few months. These authors suggest further research on the ecophysiology of these groups to answer questions regarding energy and material cycles within these systems. In addition to 16s sequences, the hzo locus was also studied. This is a gene that produces an enzyme that catalyzes the oxidation of hydrazine (rocket fuel; N2H4) to N2 gas. No mention is made of the possibilities for N2O production or consumption in this process. The gene is restricted to AnAOB only, or at least that is the inference based on the observation that hydrazine is a unique intermediate molecule of the anammox process.This study provides a useful example of the combination of qPCR and molecular-phylogenetic approaches in studying a microbiological system. Applied together, the two approaches allow the extraction of useful information regarding taxonomic diversity, both richness and evenness, among functional groups of organisms.
Lim J, Do H, Shin SG, Hwang S. 2008. Primer and probe sets for group-specific quantification of the genera Nitrosomonas and Nitrosospira using real-time PCR. Biotechnology and Bioengineering 99: 1374-1383.These authors developed precise primer and probe sets for TaqMan-based quantitative PCR to examine ammonia-oxidizing bacteria (AOB) associated with wastewater treatment facilities. These molecular tools have very low rates of false-positive and false-negative errors associated with them, and will be useful primarily to work on improving the nitrogen-removal capacity of wastewater treatment. However, AOB are nearly ubiquitous, such that these molecular tools will also be useful to a wide variety of less-directly-applied studies.These authors purchased eight strains of nitrifying AOB commonly found in water treatment plants, eight non-nitrifying bacteria also commonly found in such plants, and collected, identified, and purified seven strains of nitrifying bacteria directly from a pair of water treatment plants in operation. Using published sequences of the 16s rRNA genes of these organisms, sets of primers and probes were constructed.The development and evaluation of these primer/probe sets followed two basic procedures: first, “in silico” evaluation of potential primer and probe binding sites, and calculation of potential mismatches in various combinations. For example, a set developed for one organism may also be highly likely to amplify a related organism, reducing specificity of the assay. Second, sets were optimized for PCR conditions and trialed with the often-variable real sequences derived from culture collections or field samples. Iterating between these two processes allowed a series of final best-fit sequences to emerge, that have high specificity and low failure rates.As a template for developing very good molecular tools, this paper provides some excellent advice regarding qPCR primer development. I will be using a different system that does not include probes, simplifying some steps of this process, but the basic pattern of invention, computer evaluation, wet-lab evaluation, further computer evaluation, and refinement will still be useful.
Himmelheber DW, Thomas SH, Löffler FE, Taillefert M, Hughes JB. 2009. Microbial colonization of an in situ sediment cap and correlation to stratified redox zones. Environmental Science & Technology 43: 66-74.These authors previously studied the changes in geochemistry associated with the common practice of adding a sediment cap to cover contaminated sediments at the sediment-water interface. Such caps are commonly clean sand, with the underlying idea being the layer of sand provides a transport barrier to various contaminants moving through the system by diffusion. Sediment geochemistry, like soils, includes layers of redox conditions generated by both biotic and abiotic factors. These zones of chemical conditions migrate upwards when a sediment cap is added; not surprising considering the effect the cap has on diffusion of oxygen and other chemicals important for redox considerations.This study shows that the microbial populations also migrate upwards when a cap is added. The primary concern here seems to be the effect this population shift may have on the transport and decontamination of such pollutants as are often found in the river-bottom sediments of the eastern USA. The primary effect is likely positive: populations of bacteria and archaea in sediments will metabolize, mineralize, and generally detoxify most compounds moving up from the sediments to the cap. A few classes of contaminants, however, may not be decontaminated and it is possible their transport and release into the water column may be accelerated by these microbes.There are two key parts of the methods of this paper that interest me. First, the microbial populations were analyzed by a range of techniques including real-time quantitative PCR (qPCR). The procedure of primer design, evaluation, and data interpretation looks very similar to what I will be attempting with my own samples. Second, diversity estimates for the various strata within the sediments, derived from qPCR data, includes the use of the statistical technique Canonical Correspondence Analysis. This allows direct testing of hypotheses regarding the relationship between environmental parameters, in this case depth below surface, and estimates of biodiversity such as the Shannon-Weiner index.
Kellman L, Kavanaugh K. 2008. Nitrous oxide dynamics in managed northern forest soil profiles: is production offset by consumption? Biogeochemistry 90: 115-128.These authors measured surface flux and subsurface profiles of N2O at a number of paired sites in the managed forest of Nova Scotia. Half of the sites were clear-cut harvested three years before the study, the other half more than 50 years previously. Climate factors such as air temperatures and solar radiation were consistent across the study area. Fluxes and profiles were measured periodically through a 9-month snow-free period in 2005, from early March to late November.Surface fluxes were measured by pulling samples into evacuated containers from chambers mounted on permanent collars. Similarly, profiles were measured by sampling from permanent probes buried in the walls of soil pits. Actual measurement of gas concentrations were in the laboratory using a gas chromatograph system. The soil probes consist of 50cm PVC tubes, covered with a “water resistant porous membrane” (could they be using Gore-tex?) and buried in the walls of pits at depths of 0, 5, 20, and 35cm, with 0 at the mineral soil-organic layer interface. This provides a 50cm-long sampling space at four depths, replicated across 40 sites. The relationship between profile N2O concentrations and surface flux was almost always non-significant. These authors attribute this lack of correlation to consumption of N2O in the soil profile. In contrast, the studies that have linked CO2 profiles to surface flux have relied on the (probably true) assumption that CO2 is not consumed in the soil, and moves through diffusion in a manner that can be predicted from soil physics. N2O profiles that include regions of consumption are complicated by the biological and chemical factors that control production and consumption, as well as movement. All of this leads to a disconnection between soil N2O cycling and surface-atmosphere exchange.This paper should almost certainly be included in the introduction, methods, and/or discussion section(s) of my pits & probes manuscript. This is one of the few studies I have found that examined N2O in soil profiles; most others appear to focus on CO2 or in some cases the biogeochemistry of CH4.
Lamb EG, Cahill JF, Dale MRT. 2006. A nonlinear regression approach to test for size-dependence of competitive ability. Ecology 87: 1452-1457.These authors describe a statistical analytical technique, “nonlinear regression”, that potentially provides more information than linear regression techniques including ANCOVA. The basic linear regression formula includes the intercept and the slope as parameters; nonlinear regression adds an exponent parameter. In this paper, these parameters are referred to as k1 (intercept), k2 (slope), and k3 (exponent), in the formulay = k1 + k2x^k3
where y is the response variable and x is the explanatory variable.These authors tested their technique on three example datasets from previous studies. The first example is most thoroughly examined, and involves a plant competition experiment. One key feature of all of the example analysis is the dataset must be paired, such that each data point on the x axis corresponds to a partner data point on the y axis. In the plant competition example, individual plants that did not experience competition are paired with individuals that did, because each pair of plants was grown in a communal pot that was treated at the pot level with manipulations such as fertilizer application. The continuous dataset is plant size, measured as the absolute gain in mass over the course of the growing season.The exponent parameter k3 can take on any value between negative and positive infinity, to describe curves that may be accelerating, saturating, or straight. At values of 1 or -1, k3 is not informative as a parameter, and should be discarded from the model. This analysis is based on a model-building and model-testing technique, where models with various values for the three parameters are tested against null models and each other in an iterative fashion to find the model that best fits the data.This approach is likely to be useful in the analysis of some of the data collected at Alexandra Fjord in 2009.
Michelsen, A., Graglia, E., Schmidt, I.K., Jonasson, S., Sleep, D., and Quarmby, C. 1999. Differential responses of grass and a dwarf shrub to long-term changes in soil microbial biomass C, N and P following factorial addition of NPK fertilizer, fungicide and labile carbon to a heath. New Phytologist 143(3): 523-538.These authors measured the responses of two tundra plants, one graminoid and one shrub, to additions of fertilizer, labile carbon, and a fungicide over four years in a heath in subarctic Sweden. The underlying hypothesis is that differences in growth strategy between these two groups of plants would be reflected in their responses to changes in microbial biomass, also measured by these authors.Molecules inaccessible to plants by virtue of being incorporated into microbial cells in soils dominate their respective pools. For example, in this study, microbial N concentrations were up to 50 times higher than free, inorganic N such as ammonium. Similar ratios were found for C and P. Rapid incorporation of these nutrients into microbes therefore constitutes competition between microbes and plants. Consistent with the variety of ecological and growth strategies that plants have evolved interacting with each other and with the abiotic features, plant strategies impact their interactions with microbes and how they react to such competition.As predicted, the graminoid Festuca ovina responded rapidly to changes in nutrient levels. Added N from fertilizers led to increased ground cover by this grass, while added labile C, in the form of sugar, led to decreased ground cover, presumably due to a rapid increase in microbial biomass and associated increased microbial uptake of soil N and P. In contrast, the shrub Vaccinium uliginosum did not appreciably react to changes in nutrient levels, consistent with a long-view growth strategy and successful escape from severe competition with soil microbes. This may have been mediated by the mycorrhizal associations the shrub has but the graminoid lacks.This paper, interestingly, does not cite any of the work by Wardle or Nilsson, the authors who severely criticized the earlier work by this research group (Michelsen et al. 1995).
Wardle DA, Nilsson M-C. 1997. Microbe-plant competition, allelopathy and arctic plants. Oecologia 109: 291-293.These authors critique Michelsen et al. (1995), a study that came to several important conclusions regarding the interactions between Arctic plants and the soil microbial communities. This is a very negative review of that paper, in which these authors question almost all of the conclusions of Michelsen et al. (1995).These authors make two main criticisms. First, they question the measures of soil microbial activity used by the earlier paper. Second, they question the conclusions regarding the allelopathy of Empetrum hermaphroditum. Soil microbial activity was measured by Michelsen et al. (1995) in two ways: soil respiration, and soil ergosterol content. Neither approach is necessarily informative about one of Michelsen et al.’s (1995) main claims, that soil microbial biomass was increased by the addition of plant leaf extracts. There are a number of studies, many of them with Nilsson as a co-author, in which a lack of association between microbial biomass and soil respiration was demonstrated. Furthermore, ergosterol is presented by Michelsen et al. (1995) as an indicator of fungal biomass, but previous work by Newell and colleagues (e.g. Newell and Fallon, 1991; Newell 1992) showed that ergosterol is not a reliable indicator of biomass nor is it useful as a proxy measure of soil fungal activity; the ratio of ergosterol to fungal biomass is highly variable. From the reference list in this short paper, it appears that Wardle and Nilsson had, by early 1997, completed a considerable body of work regarding the allelopathic and other ecological interactions of E. hermaphroditum in sub-arctic environments. The conclusion by Michelsen et al. (1995) that the chemicals released by this plant have a greater impact on microbial communities than potential surface-plant competitors is not supported by this work by Wardle, Nilsson, and their colleagues.The conclusion of Michelsen et al. (1995) that currently has the most direct bearing on my own work is that key plant traits often possessed by prostrate shrubs in tundra ecosystems such as a high root: shoot ratio and storage of nutrients such as nitrogen in the roots allow those plants to escape from or outcompete soil microorganisms. This conclusion was not addressed by these authors, but given the devastation inflicted upon the other conclusions, my confidence in the utility of Michelsen et al. (1995) in addressing issues of interactions between Cassiope tetragonal and soil microbial communities has been shaken.
Michelsen A, Schmidt IK, Jonasson S, Dighton J, Jones HE, Callaghan TV. Inhibition of growth, and effects on nutrient uptake of arctic graminoids by leaf extracts – allelopathy or resource competition between plants and microbes? Oecologia 103: 407-418.These authors conducted an experiment to examine the potential allelopathic effects of plant leaf extracts among a few species of arctic tundra plants. Three arctic/subarctic plants suspected of releasing phytotoxic compounds upon their competitors including Cassiope tetragonal were harvested and their leaves and branches ground up to make leaf extracts. Three species of arctic graminoids (Carex bigelowii, Festuca vivipara, Luzula arcuata) were then treated with these extracts while growing in either sterilized or non-sterilized soil in greenhouses in southern Sweden. All of the graminoids, the soil they grew in, and two of the leaf extract-providing plants came from a montaine subarctic hillside in northern Sweden; the third leaf extract came from Betula pubescens ssp. tortuosa, a birch, were collected from individuals growing near the tree line at 450m altitude near Abisko Scientific Research Station, above the Arctic Circle in a subarctic ecosystem.The experimental design was factorial, with two soil types (sterilized vs. non-sterilized) and four leaf-extract treatments including a control of distilled water. In addition to growth of the graminoids, measurements were made of the chemicals in the leaf extracts and soils, nutrient uptake by excised roots, soil ergosterol content, and soil respiration. Excised roots take up nutrients in a manner directly correlated to the nutrient-limitation status of the plant; more phosphorus-starved plants, for example, have roots that more rapidly take up phosphorus when offered. Soil ergosterol content is a measure of fungal biomass, while soil respiration was taken as a measure of total microbial activity.Sterilized soil had higher extractable nutrients, probably as a result of the breakdown of microbial cells during autoclaving. Nitrate levels were negligible, both in soils and in leaf extracts; nitrate was a component of the dilute nutrient water used to maintain the plants while growing. Some mycorrhizae were found, but they covered less than 1% of the roots in non-sterilized soil, and had no impact on other measured parameters.The highest growth of all three test graminoids was recorded in sterilized soil with no extract added (i.e. distilled water added instead of leaf extract solution). Plants growing under these conditions experienced no inhibition from the materials of other plants, and did not compete with soil microbes for nutrients, at least during the early stages of the experiment before microbes recolonized the sterilized soils. Recolonization was much faster by prokaryotes than by fungi, as measured by the contrast in soil respiration rates and soil ergosterol contents. Recolonization also varied between leaf extract treatments, with a negative correlation between microbial activity and plant growth; strongly growing plants were able to outcompete colonizing microbes, while poorly growing plants were further inhibited by colonizing and rapidly growing microbes.All three leaf extracts significantly reduced growth of all three graminoid species. However, it is not clear that allelopathy alone was responsible for this effect. The results of this study indicate that competition between plants and microbes also played a major role. In particular, the components of the leaf extracts, especially labile carbon and (in the case of the Betula extract) phosphorus, appeared to stimulate the microbial community, increasing competitive pressure on the plants. Added nitrogen, for example, appears not to have much benefited the plants, as their roots were N-limited when grown in non-sterilized soil even though the leaf extracts included high concentrations of inorganic nitrogen.The susceptibility of plants to this combined effect of allelopathy and microbial competition probably varies by species. Plant traits of particular importance are probably the root: shoot ratio, in which plants with more robust roots are less harmed, and the storage of nutrients such as nitrogen in plant roots, in which plants with a growth strategy favouring nutrient storage rather than immediate use are less harmed. Such traits appear to be widespread among the dominant plants of the Arctic tundra, including Cassiope tetragonal and probably other prostrate shrubs. Many of these plants may form associations with mycorrhizal fungi, which provide some protection against microbial competition.This paper is directly relevant to the discussion section of my current-high-priority Pits & Probes manuscript. The patterns of soil respiration and microbial GHG activity under some of the lowland communities are consistent with successful competition against soil microbes by Cassiope tetragonal and possibly Salix arctica plant roots.
This paper was critiqued quite harshly by Wardle and Nilsson (1997).


